And It's About Time There Was Some Support For Cushing's!
![]()
![]()
From Children's Hospital Informatics Program (CHIP)
ABSTRACT
This paper describes the construction of a computer model that simulates the hypothalamic-pituitary-adrenal axis (HPA axis) regulation of cortisol production. It is presented to illustrate the process of physiological modeling using standard "off the shelf" technologies. The model simulates components of the HPA axis involved in the continuous secretion and elimination of cortisol, adrenocorticotropin (ACTH), and corticotropin releasing hormone (CRH). The physiological relations of these component pieces were modeled based on the current knowledge of their functioning. Rate constants, half lives, and receptor affinities were assigned values derived from the experimental literature. At its current level of development the model is able to accurately simulate the timing, magnitude and decay of the ACTH and cortisol concentration peaks resulting from the ovine-CRH stimulation test in normal and hypercortisolemic patients. The model will be used to predict the effects of lesions in different components of the HPA axis on the time course of cortisol and ACTH levels. We plan to use the model to explore the experimental conditions required to distinguish mechanisms underlying various disorders of the HPA axis, particularly depression. Efforts are currently underway to validate the model for a large variety of normal and pathological perturbations of the HPA axis.
HPA axis dysfunction has been found to occur during psychiatric illness [1-5]. Therefore, study of HPA axis functioning is important to further our understanding of the underlying pathophysiology of psychiatric disorders.
The HPA regulation of cortisol is accomplished through a complex network of interacting components. Despite increasingly detailed knowledge about each of these components, it remains difficult to predict from this information how the network will perform.
One possible approach to understanding network functioning is to build a simulation that allows us to model, quantitatively, the functioning of these components within the context of the network (HPA axis). This tool would make it possible to subject hypotheses about the HPA axis to more rigorous tests and would provide a platform on which experiments could be planned to decide between competing hypothesis [6]
Two prior attempts at modeling systems of hormone regulation demonstrate two possible approaches to this task. Sharma and Gabrilove [7] created a model of the HPA axis by simulating each enzymatic step in the synthesis of cortisol. The difficulty with this approach lies in the degree of mechanistic detail required to simulate the system. Seventy five independent parameters were used, many of which had not determined experimentally, making the utility of the model difficult to assess. Bing-Zhen and Gou-Min [8] approached the modeling task in an alternative fashion by using a set of differential equations that fit the time course of known hormonal oscillations. They acknowledge, however, that there is insufficient experimental data to fully specify their equations. Therefore, they fit their equations to the empirically observed time courses of hormone levels. While these equations give a satisfactory phenomenological description of the system, they are of limited usefulness in furthering understanding of the molecular mechanisms of the axis as the form of the differential equations is largely independent of known mechanisms for the production of these hormones.
This paper describes a simulation of the HPA Axis using an approach that takes a middle road between these two prior attempts at modeling hormone regulation. It seeks to simulate only those mechanistic details that are experimentally observable in order for the model to be relevant to those conducting experiments on HPA axis functioning.
Background:
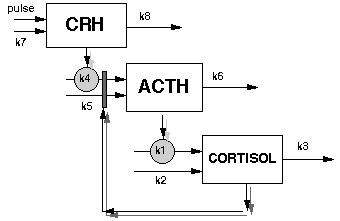
To provide a background to the simulations described in this paper, the physiologic interactions of the HPA axis are reviewed briefly. CRH and arginine-vasopressin (AVP) produced in the hypothalamus stimulates the pituitary to secrete ACTH. ACTH stimulates release of cortisol from the adrenal gland. Cortisol exerts a negative feedback directly on the pituitary and also on the synthesis and secretion of CRH and AVP. The secretion and effects of CRH and AVP are influenced by neuro-transmitters in the hypothalamus and deeper in the CNS as well as circulating epinephrine [9].
All simulations were developed and run on a Macintosh SE/30, with a 68030 processor running at a speed of 25 mHz. EXTEND (Imagine That, Inc., San Jose, CA) a commercially available simulation modeling package was used for building the simulations.
Simulation Construction
For this initial experiment, the granularity of mechanistic detail was chosen that would be sufficient to simulate the time course of ACTH and cortisol plasma levels observed during a standard test of HPA functioning, the Ovine-CRH stimulation test (oCRH stimulation test). The test is performed by administering 1 ug/kg body weight to patients at a specified time of day, then assaying plasma cortisol and ACTH levels for at least 2 hours. The simulations were built in modules, each module representing the synthesis and degradation of a single hormone (cortisol, ACTH, CRH). The following are the differential equations used in each of the modules:
Cortisol module: Eq. (1)
d[cortisol]/dt = k1*[ACTH] + k2 - k3*[cortisol]
ACTH module: Eq. (2)
d[ACTH]/dt = k4*[CRH]+k5 - k6*[ACTH]+ (k4*[CRH]+k5) *Imax*[cortisol]/(Kd+[cortisol])
CRH module: Eq. (3)
d[CRH]/dt = pulse function,f(t) + k7 - k8[CRH]
Where [X] is concentration of X. kn is a rate constant. Imax is the saturation level of inhibition. Kd is the disociation constant for the glococorticoid II receptor.
A first order nonsaturating function was chosen for each of the production and degradation terms in order to simplify the model for these initial experiments. A saturating function was needed, however, for the term in Equation 2 that represents the inhibition by cortisol at the level of the pituitary of ACTH release. Attempts to model this inhibition with a first order inhibition constant failed. When any constant was used that could produce adequate inhibition within the first twenty minutes following simulated oCRH stimulation, negative values for the ACTH level were produced. This result suggested that there must be saturation of the inhibition mechanism at physiologic levels of cortisol. This prediction is confirmed by the experimental literature's finding that cortisol's maximum capacity to inhibit ACTH production is achieved before cortisol reaches it's highest physiologic plasma levels [10,11]. As there is empirical evidence that the glucocorticoid type II receptor mediates the inhibition of ACTH secretion that occurs through the direct action on the pituitary [11], a function relating inhibition to occupancy of the glucocorticoid type II receptor was used.
The modules were built sequentially. First the module simulating the production and degradation of cortisol was constructed. This was done by first embedding Equation 1 into the program and using the Euler method for integration of the cortisol level. A cycle time representing one minute of simulated time was chosen because it provided stable performance while still allowing the simulation to be completed on the Macintosh within a minute of real time. To simplify the model, cortisol was assumed to be diluted in a single 4000 ml plasma compartment. Plasma protein binding of cortisol and other hormones was not included in the model. The dilution was assumed to be complete within the one minute cycle time. The degradation constant (k3) was estimated from the published half life of cortisol in plasma [12]. The stimulation constant, k1, was set empirically using the literature regarding the oCRH stimulation test. An initial estimate of the constant was made by using Equation 1. The initial value was estimated based upon the ratio of peak change in plasma ACTH to peak change change in plasma cortisol values during the oCRH stimulation. test. This value was then varied in test simulations of the module until a value of k1 was found that produced the published magnitude of the cortisol peak following a simulated pulse of ACTH. The magnitude of the ACTH pulse simulated actual values of ACTH production during oCRH stimulation.
The same process was repeated for the module simulating the secretion and degradation of ACTH. The CRH stimulation term, k4, was based upon the expected rise in portal CRH and the observed rise in ACTH plasma levels during the oCRH stimulation test [12]. The term representing the inhibition of the pituitary by cortisol was left out of this module until later in the model building process, when a separate module was built for it and attached to the ACTH module. The ACTH module was connected to the cortisol module to allow the simulated plasma ACTH concentration derived from the ACTH module to set the value of the ACTH concentration for the Cortisol module.
In the same way the CRH module was built. A pulse function that simulates the presence of 50 micrograms of oCRH in the plasma volume was used to initiate the oCRH stimulation test simulation. The integrated CRH level was used to set the CRH concentration for the ACTH module.
TABLE 1: Constant Values
Simulation 1: Normal Simulation 2: Simulation 3,
Acute Adrenal Adenoma
Hypercortisolemia Chronic Hypercortisolemia
Equation k1 = 5x105 same as same as simulation 1
1 simulation 1
k2 = 0.01 (mcg/min) k2 k2 = 15
=15(micrograms/min (micrograms/min)
)
k3 = 0.01 same as same as simulation 1
simulation 1
Equation k4 = 10 same as k4 = 1.0
2: simulation 1
k5 = 4x10-3(mcg/min) " same as simulation 1
k6 = 0.035 " "
Imax = 0.99 " "
Kd = 0.004 (mcg/ml) " "
Equation pulse CRH = 50 mcg over same as same as simulation 1
3: 1 min simulation 1
k7 = 1x10-6(mcgs/min) " "
k8 = 0.01 " "
The cortisol inhibition of ACTH secretion by the pituitary was added to the model after the above three modules had been connected. The inhibition function used is shown in Equation 2. Glucocorticoid inhibition effects are very complex [13]. In order to simplify the model for these initial experiments only fast inhibition at the pituitary was modeled. The Kd was set at value defined for the type II glucocorticoid receptor (converted to micrograms/ml of cortisol) [14]. The Imax was set at .99 to simulate the effect of very strong fast inhibition at the level of the pituitary on the oCRH stimulation test . The Inhibition module was connected in order to decrease ACTH production prior to its numerical integration into the system.
Once the above modules were connected, the baseline production constants, k2, k5, and k7, were set empirically so that the correct baseline ACTH and cortisol plasma concentrations were achieved by the simulation prior to the simulated administration of oCRH [12].
Numerical Integration
During the simulations these equation modules were updated sequentially in the "downstream" direction from CRH to ACTH to Cortisol (each cycle representing one minute of simulation time). The Euler method was used for numerical integration of the equation for each hormone and the concentration of ACTH and cortisol was updated with each cycle. Simulations were run for 500 minutes prior to the simulated oCRH stimulation test in order for baseline Cortisol and ACTH levels to reach known baseline equilibrium values [12]. At 500 minutes, the simulated administration of the oCRH occurred and the simulation continued for an additional 120 minutes of simulation time.
The simulations were truncated after 120 minutes of simulation time for two reasons. 1) This is the usual duration of the oCRH stimulation test, and 2) a secondary peak of ACTH and cortisol occurs beyond 120 minutes due to the long half life of oCRH and production of new ACTH from pro-opiomelanocortin [15]. Simulation of this second peak was not attempted at this stage of the development of this tool, although it is the subject of planned future work.
RESULTS
For these simulations, the ovine-CRH stimulation test in normal controls and in hypercortisolemic patients was used as a model.
The results of the simulations were compared to published results in normal controls and patients with adrenal adenoma and hypercortisolemia [12, 16]
The constants used for the simulations are listed in Table 1. The simulation results for the normal oCRH stimulation test are shown in figure 1. The magnitude and timing of the cortisol and ACTH peaks are within published ranges of normal values for this test [12]
The initial simulation for hypercortisolemia assessed the effect of acute exposure to high cortisol levels on the results of the oCRH stimulation test (Figure 2). For this simulation the constant for baseline cortisol production, independent of ACTH stimulation, was increased. It is of note that despite an Imax set for .99, the ACTH response, although greatly diminished from normal values, is higher than what would be found with chronic hypercortisolemia. This is consistent with the observation that the ACTH response to many stressors cannot be completely inhibited by acute exposure to high cortisol levels [10, 11].
FIGURE 1: Simulation I:
Normal oCRH Stimulation TEST
FIGURE 2: Simulation II:
Acute Hpercortisolemia
3: Simulation III
Chronic Hypercortisolemia
In order to successfully simulate the oCRH induced ACTH response in patients with chronic hypercortisolemia, the suppressive effects of prolonged hypercortisolemia had to be taken into account. For this simulation (Figure 3), in addition to increasing the baseline cortisol production constant, the constant which represents CRH stimulation of ACTH production was decreased by an order of magnitude. This additional step was taken to simulate the profound suppressive effects on corticotrophs during chronic feedback inhibition of the HPA axis. This is consistent with the finding that ACTH response to CRH remains depressed long after hypercortisolemia has been treated by surgical removal of an adrenal adenoma [16]
The purpose of this paper is to describe a process of constructing a working simulation of HPA axis regulation of cortisol production and to report initial results obtained using this technique. The program was able to successfully simulate 1) the plasma half lives of both of these hormones and 2) the magnitude and timing of the of cortisol and ACTH peaks in response to an oCRH stimulation test in normal individuals and patients with hypercortisolemia .
There are several possible ways to proceed in further building this tool:
1) Greater mechanistic detail can added to the model, by integrating knowledge of the molecular events involved at a cellular level and/or by extending the model deeper into the central nervous system.
2) Maintain the current level of detail of the model, but attempt better estimates of the constants by requiring them to fit the results of additional experiments (i.e.. different diagnostic tests of the HPA axis). Monte Carlo simulations can be used to define ranges of values for the constants that are consistent with experimental results.
3) Since the process of adding complexity can increase error, an approach combining 1 and 2 above can be taken. That is, the level of mechanistic detail simulated will vary in different parts of the model to another and will also depend on the questions being asked.
4) Generate a range of models and selecting the minimalor optimal model by applying formal criteria [6].
This paper illustrates an approach, implemented with "off the shelf" software, to create simulations that closely follow clinical and experimental observations. A central feature of our effort has been to ground our model with mechanistic knowledge that is well supported by the experimental literature without adding detail that is unnecessary or untestable. After rigorous iterative testing and validation, the predictions of HPA system behavior by this model can serve as guides to further basic and clinical research.
REFERENCES
[1] L. Musselman and C. B. Nemeroff . The Role of Corticotropin Releasing Factor in the Pathophysiology of Psychiatric Disorders. Psychiatric Annals, 23: 676 - 681.1993.
[2] J. Rothschild. The Dexamethasone Suppression Test in Psychiatric Disorders. Psychiatric Annals, 23: 662-670.1993.
[3] R. R. Krishnan. Pituitary and Adrenal Changes in Depression. Psychiatric Annals, 23: 671-675.1993.
[4] E. P. Murphy and O. M. Wolkowitz. The Pathophysiologic Significance of Hyperadrenocorticism: Antiglucocorticoid Strategies. Psychiatric Annals, 23: 682 -690.1993.
[5] N. Starkman. The HPA Axis and Psychopathology: Cushing's Syndrome. Psychiatric Annals, 23: 691-701.1993.
[6] Carson, C. Cobelli and L. Finklestein. The Mathematical Modeling of Metabolic and Endocrine Systems.John Wiley and Sons. New York. 1983.
[7] C. Sharma and J. L. Gabrilove. A Study of the Adrenocortical Disorders Related to the Biosynthesis and Regulation of Steriod Hormones and their Computer Simulation. Mount Sinai Journal of Medicine, 42: S2-S39.1975.
[8] Bing-Zheng and D. Gou-Min. An Improved Mathematical Model of Hormone Secretion in the Hypothalamo-Pituitary-Gonadal Axiin Man. Journal of Theoretical Biology, 150: 51-58.1991.
[9] B. Martin and S. Reichlin. Clinical Neuroendocrinology.F. A. Davis. Philadelphia. 1987.
[10] Dayanithi and F. A. Antoni. Rapid as well as Delayed Inhibitory Effects of Glucocorticoid Hormones on Pituitary Adrenocorticotropic Hormone Release are Mediated by Type II Glucocorticoid Receptors and Require Newly Synthesized Messenger Ribonucleic Acid as well as Protein. 1989.
[11] E. McCutcheon and E. H. Oldfield . Cortisol: Regulation, Disorders, and Clinical Evaluation. In: Neuroendocrinology , ed. D. L. Barrow and W. R. Selman. Williams and Wilkins, Baltimore, 1992.
[12] T. Jones and B. Gilham. Factors Involved in the Regulation of Adrenocorticotropic Hormone/beta-Lipotropic Hormone. Physiological Reviews, 68: 743-818.1988.
[13] M. H. M. Reul and E. R. de Kloet. Two Receptor Systems forCorticosterone in the Rat Brain: Microdistribution and DifferentialOccupation. Endocrinology, 117: 2505-2511.1985.
[14] R. Debold, R. V. Jackson, T. C. Kamilares, W. R. Sheldon, G. S. Decherney,D. P. Island and D. N. Orth. Effects of Ovine Corticotropin-ReleasingHormone on Adrenocorticotropin Secretion in the Absence of GlucocorticoidFeedback Inhibition in Man. Journal of Clinical Endocrinology andMetabolism, 68: 431.1989.
[15] Fukata, T. Usui, T. Tsukada, N. Y., T. Koh, T. Ishihara, I. Tanaka, K.Uchida, T. Yoshimasa, R. Nakano, M. Nishikawa and H. Imura. Effects ofRepetitive Administration of Corticotropin-Releasing Hormone Combined withLysine Vasopressin on Plasma Adrenocorticotropin and Cortisol Levels inSecondary Adrenocortical Insufficiency. Journal of ClinicalEndocrinology and Metabolism, 71: 1624-1631.1990.